|
|
||
09/07/10 |
|
|
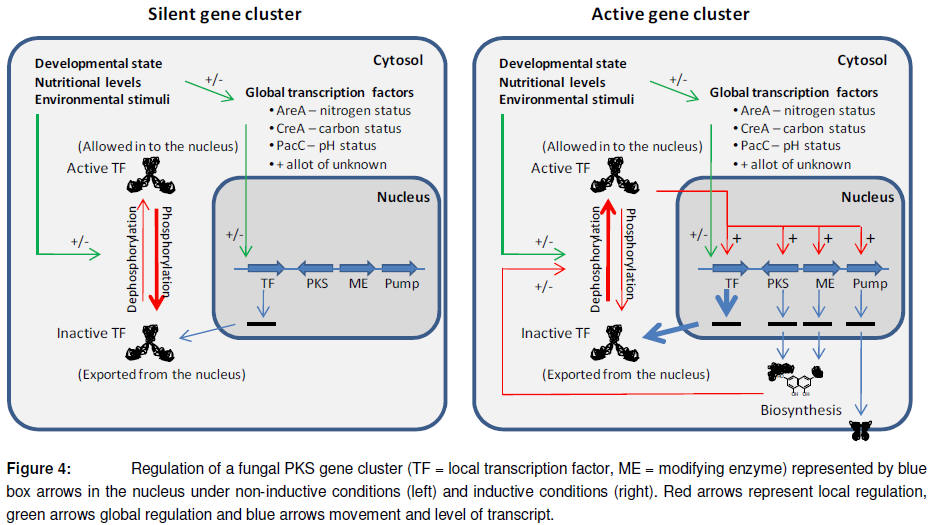
The first seven decades of polyketide research was led by chemists and biochemists, and little attention was given to the enzymes responsible for catalyzing the required reactions. The first breakthrough on this front came in 1971 where Dimroth and co-workers succeeded in preparing a cell free extract from Penicillium patulum that retained the ability to synthesize 6-methylsalicylic acid (Dimroth et al., 1971). Experiments using this system led to an understanding of the basic concepts of polyketide biosynthesis and the polyketide synthase. The next significant recognition came with the advance of molecular biology in the 1980s, which allowed for the direct linking of polyketide biosynthesis to the responsible genes. The subsequent period was dominated by the study of polyketide biosynthesis in Streptomycete bacteria and its genetic foundation, and included work on what is now known as type I modular polyketide synthases and type 2 polyketide synthases (reviewed by (Hopwood, 1997)). Analysis of the genetic basis for polyketide synthesis in bacteria showed that entire biosynthetic pathways typically were encoded by clusters of co-regulated genes (In addition to the typical clustering due to the operon structure utilized by bacteria). The first fungal polyketide synthase to be cloned was 6-MSA synthase (6-MSAS) from Penicillium patulum (Beck et al., 1990). Sequence analysis of the 6-MSA synthase proved that it was very different from the PKS system described in bacterial systems and that it showed a higher degree of similarity to mammalian fatty acid synthases (O'Hagan, 1991). The majority of fungal PKSs described to date are classified as iterative Type I PKSs (iPKSs), however in 2005 Seshime and co-workers reported the first finding of a Type III PKS (chalcone synthase (CHS)-like) in Aspergillus oryzae (Seshime et al., 2005).Similarly to the findings in bacterial systems, fungal genes encoding enzymes for a common biosynthetic pathway also show co-regulation and clustering in the genome (Keller and Hohn, 1997). However, unlike their bacterial counterparts fungi do not have the luxury of operon and polycistronic mRNA, and the individual genes are regulated by individual or bidirectional promoters. Expression of the clusters is typically controlled by a local trans-acting transcription factor which is also encoded by the gene cluster, creating a positive feedback system (Figure 4).
The expression and activity of the local transcription factor is in turn regulated by global regulators (Including both transcription factor systems controlling expression of the local transcription factor and enzymes that control the subcellular localization of the local transcription factor protein, typically by phosphorylation and de-phosphorylation) ensuring that the level of polyketide synthesis is adapted to the environment, nutritional status, developmental state and overall needs of the fungus (Calvo et al., 2002). It has been proposed that secondary metabolism is regulated by a single on/off switch, possibly by histone/chromatin remodelling (Kale et al., 2007; Kale et al., 2008), but so far the theory is supported by very limited experimental data. The best described complex fungal polyketide biosynthetic system to date is the carcinogenic aflatoxin and the related stigmatocystein systems in Aspergillus flavus and A. paraciticus. Work in the late 1990’s and early 2000’s has provided an almost complete understanding of the 28 required biosynthetic steps as well as an understanding of the complex regulation, including the effect of genomic organisation (reviewed by (Yabe and Nakajima, 2004), (Cary and Ehrlich, 2006) and (Bhatnagar et al., 2006)).
Examples of fungal secondary metabolite gene clusters in Fusarium and Aspergillus
References:
|
|||||||||||||||||||||||||||||||||||
Dette sted blev sidst opdateret 09. July 2010