|
|
||
09/07/10 |
|
|
The best described fungal polyketide gene clusterThe gene clusters responsible for the biosynthesis of polyketides in fungi typically consist of a PKS gene, genes encoding hydrogenases, oxidases, transporters and regulatory genes (Keller & Hohn 1997) (Yu et al. 1996). Examples includes aflatoxin and sterigmatocystin (Bhatnagar et al. 2003), AK-toxin (Tanaka et al. 1999), fumonisins (Bojja et al. 2004), HC-toxin (Ahn & Walton 1999), lovastatin (Kennedy et al. 1999), melanins (Tsai et al. 1999) and paxilline (Young et al. 2001). The currently best-described polyketide gene cluster, with respect to regulation, is the aflatoxin (AF) gene cluster known from Aspergillus parasiticus, A. oryzae, A. sojae, A. nomius, A. pseudotamarii, A. bombycis and A. ochraceoroseus and the closely related sterigmatocystin (ST) gene cluster known from A. nidulans, A. versicolor, A. ustus and some A. oryzae strains (Woloshok et al. 1994). Research in the genetic basis of AF production started the 1960, provoked by the death of over 100,000 turkeys in England, all within a single production season. All the affected animals had been feed with Aspergillus contaminated cotton seeds. The deadly metabolite was identified and named aflatoxin. This polyketide group belongs to the furanocoumarins (Deacon 1997) (Woloshok et al. 1994). AF’s can be divided in to two main groups (B and G), based on their differences in fluorescence emission spectrums (B = blue and G = green) (Deacon 1997). In addition to being acutely toxic in high concentrations, both classes has been shown to be mutagenic, teratogenic, hepatocarcinogenic and immunosuppressive (Eaton & Groopman 1994). In the initial days of molecular biology their strong mutagenic properties was utilized to generate random mutations in organisms of interest (Griffiths et al. 1999). Aflatoxin continues to be a serious problem in development countries, where no efficient control strategies exist apart from testing every individual harvest batch (Calvo et al. 2004).
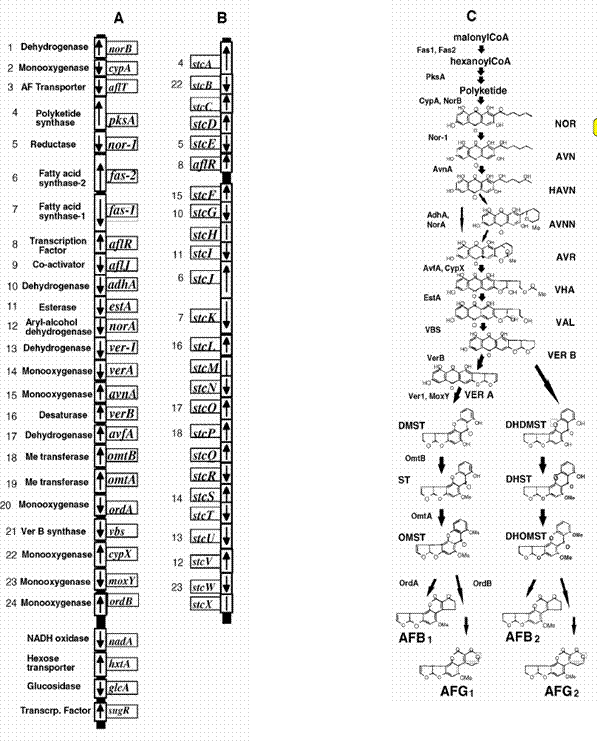
Genetic basis for the production of aflatoxinUntil now, all the genes that have been shown to be involved in the biosynthesis of AF are located within a 70 kb large region of the A. flavus and A. parasiticus genomes (Figure 1). These identical gene clusters consists of 24 genes of which 20 are involved directly in the biosynthesis of AF by catalyzing one or more steps. The remaining 4 genes are transcriptional regulators and AF-pumps (Flahery & Payne 1996). In 1996, Brown et al. showed that A. nidulans also has the PKS (pksA) responsible for synthesis of the precursor AF polyketide in A. flavus and A. parasiticus. However, A. nidulans does not produce AF, but ST, a known intermediate from the AF biosynthesis pathway (Brown et al. 1996). Later analysis of the ST gene cluster has shown that it shares 16 genes with the AF gene cluster and contains an additional 9 genes, however restriction enzyme mapping has shown that the shared genes are arranged differently in the ST and AF gene clusters (Brown et al. 1996) (compare figure 1A and B).
Regulation of aflatoxin production The AF production is regulated both at the transcriptional and post-translational levels (Flaherty et al. 1996), but the transcriptional regulation has received most attention. In 1993 Chang et al. identified the pathway specific transcription factor aflR (initially named afl-1 and apa-2) in A. parasiticus within the AF gene cluster. Bioinformatic analysis of AflR has revealed that it belongs to the “binuclear zinc cluster” superfamily of transcription factors, which typically binds to their DNA targets as homodimers (Bhatnagar et al. 2003). Deletion of aflR results in non-aflatoxigentic mutants, due to reduced expression of the genes found in the AF gene cluster. AflR functions as a positive acting transcription factor of the AF gene cluster (Takahashi et al. 2002). AflR orthologs has later been identified in all other AF and ST producing Aspergillus sp. (Chang et al. 1993). Foot-printing analysis with promoter DNA from the affected genes and recombinant AflR protein has led to the identification of the DNA target; 5’-TTAGGCCTAA-3’ in A. flavus and A. parasiticus and 5’-TCGN5CGA-3’ in A. nidulans (Sidhu 2002). Later electro mobility shift assays (EMSA) has revealed that the promoters of the following genes pose these motifs; aflR, ver1, norA, vbs, ordA, omtA, pksA, adhA, ord1, avnA, stcS, stcT, stcU and stcV (Ehrlich et al. 1999) (Cary et al. 2000).
Figure 1 (A) Aflatoxin gene clusters in A. flavus and A. parasiticus and (B) sterigmatocystin cluster in A. nidulans. (C) Shows the enzymatic steps responsible for the production of AF and ST. The numbers in A and B indicate ortholog genes in the two clusters. Illustration taken from (Bhatnagar et al. 2003).
In 1998 Meyers and co-workers identified the gene aflJ and proposed that it functioned as a pump responsible for the movement of intermediates between the different cellular compartments during the AF biosynthesis (Meyers et al. 1998). Later studies by Chang et al. (2001) have shown that AflJ functions as a co-activator to AflR. Deletion of aflJ results in a non-aflatoxigentic phenotype due to depression in the expression levels of genes regulated by AflR, except for pksA (Meyers et al. 1998). Two-hybrid analysis in S. cerevisiae with aflJ and aflR has shown that the two corresponding proteins interact in vivo (Chang 2003). These results also explain why A. sojae, which contains all the structural genes, is non-aflatogentic as it has a truncated aflR allele, which affect the interaction between the two proteins (Chang 2004). The mechanism by which AflR/AflJ affects their target genes is yet unknown, but relocation of one of their known targets, the ver-1 gene, from the AF gene cluster to the nirD locus affects its expression negatively (Liang et al. 1997). These results have been interpreted to mean that AflR/AurJ do not directly affect the target gene (by attracting the transcription machinery), but rather affects the local chromatin structure. Chang (2004) speculates that AflJ could participate in chromatin remodeling. Multiple DNA targets, as found in the AF gene cluster, should result in amplification of the effect. This hypothesis could also explain the existence of gene clusters, as this arrangement will allow for additive co-regulation of the genes found in the cluster. This hypothesis also works in evolutionary perspective as clustering and thereby co-regulation will result in expression of all the necessary genes simultaneously (Keller & Hohn 1997). The aflR promoter contains a AflR binding motif, resulting in a positive feedback system, known as autoregulation (Chang et al. 1995). Autoregulation results in an explosive expression of aflR upon induction, which in turn leads to instantaneous high expression of the genes found in the AF gene cluster. aflR and aflJ shares a bidirectional promoter (Figure 1) but only the expression of aflR is affected by binding of AflR. The autoregulation scheme presents serious problems when induction needs to be stopped, two factors has been identified which might work as off-switches: (1) the phosphorylation state of AflR (Shimizu & Keller 2001) (Shimizu et al. 2003) and (2) the existence of an yet uncharacterized repressor factor (Ehrlich et al. 1999). The transport of AflR into the cell nucleus is controlled by its phosphorylation stage, as described for other transcription factors (Lodish et al. 2003). The Aspergillus sp. Protein kinase A (PkaA) has been shown to phosphorylate AflR in vitro (Shimizu et al. 2003). A G-protein signalling pathway, consisting of FlbA, FadA (a-subunit) and adenyl cyclase, controls the activity of PkaA. The active FadA (GTP-form) activates the Adenyl cyclase, resulting in increased levels of cellular cAMP. As PkaA is a cAMP dependent kinase, this leads to its activation and phosphorylation of AflR. The phosphorylated AflR-form is either restricted from entering the cell nucleus or results in increased export from the nucleus. AflR’s exclusion from the nucleus prevents it from inducing the genes found in the aflatoxin gene cluster. The active FadA (GTP-form) can be inactivated by FlbA a RGS containing protein (Regulation of G protein Signalling) by external signals. This leads to lower cAMP levels and inactivation of PkaA, which in turn allows AflR to enter the cell nucleus and induce the aflatoxin related genes (Shimizu & Keller 2001). Cary et al. (2000) have shown that nuclear extracts from A. parasiticus contains an unknown transcription factor that can bind to the aflR promoter from position -103 to -90, but the effects of this protein is not known. Truncation analysis of the aflR promoter region has indirectly proven the existence of a yet unidentified repressor factor, as deletion of the region -280 to -118 results in increased expression of aflR and production of aflatoxin (Chang et al. 1999).
Global regulation of the aflatoxin production:The aflatoxin production is influenced by the growth conditions (N and C-source, pH of medium) and cell cycle. The effects of pH, N- and C-source have all been linked to the activity of global transcription factors, PacC, AreA and CreA respectively. All three are believed to regulate the AF production by controlling the expression level of aflR and aflJ. PacC (active when the medium is alkaline) inhibits ST/AF production, while AreA (active under N-starvations) promotes ST/AF production (Keller et al. 1997) (Ehrlich et al. 1999). The exact signalling pathways for these global transcription factors are only partly known. Sequence analysis of the Aspergillus sp. pksA promoter has shown that it has several CreA binding sites, but the effects of these has not been investigated experimentally (Ehrlich et al. 2002). The biosynthesis of AF is induced by the presence of simple carbohydrates in the medium, such as glucose, lactose and sucrose. Whether this is the direct result of CreA activity or due to activation of a hexose utilization gene cluster adjacent to the AF gene cluster, resulting in changes in the local chromatin structure, is unknown (Yu et al. 2000). Evidence of a linkage between the production of secondary metabolites and the asexual sporulation in fungi has existed for a long time. Mutations affecting the production of aflatoxin in many cases also has a negative effect on the production of conidia (both number and size are affected). A direct link was proven by Shimizu et al. (2003), who described a G-protein signalling system, consisting of FadA, FlbA and PkaA, which affects both AF production and the activity of BrlA, a conidia specific transcription factor (Shimizu & Keller 2001). Sequence analysis of the pksA promoter has shown that it in addition to AflR target sequences also contains BrlA binding motifs. Deletion of this BrlA motif has a negative effect on the AF production (Ehrlich et al. 2002). Other factors (veA, laeA and rcoA) have also been shown to influence the production of AF. Deletion of any of these three genes results in mutants incapable of producing AF. The activity of LaeA affects the production of secondary metabolites in general (Bok & Keller 2004). VeA affects the production of AF but not the development of conidia (Calvo et al. 2004). RcoA affects both AF production and conidiation (Hicks et al. 2001). Even for this well characterized biosynthesis pathway the overall regulation is still poorly understood.
References |
Dette sted blev sidst opdateret 09. July 2010